


植物に今起きている進化をつかまえろ!
田中 健太先生
進化の原動力は、なんでしょう?生物の「進化しやすさ」を決めているものとは?アブラナの一種である「ミヤマハタザオ」の研究から、進化を加速するメカニズムのひとつが見えてきました。それは、同じ遺伝情報を余分に持つことで生まれた「進化を可能にするゆとり」。標高30mから3000mまでの多様な環境に適応するべく、ミヤマハタザオは今まさに進化中です。田中健太准教授は、野外での生態調査と遺伝子レベルでの解析を組み合わせて、今そこで起こっている進化に迫っています。
進化のブラックボックスに斬りこむ
進化を研究するとは一体、どういうことなのでしょうか?田中先生に聞いてみると、「世の中の進化のパターンって、不思議なことがいっぱいあるんだよ。スゲやアザミの仲間とか、あるものは爆発的な多様化をして1属400種にも増えている一方で、1科1属1種も全然珍しくない。あるグループはすごく進化して多様化しているのに、あるグループは全然多様化していない。それはなんでなの?今は、こんな単純な質問にも誰も答えられないんだよ。」と熱い言葉が返ってきました。
ダーウィンの進化論はご存知ですか?昔々、サバンナで暮らすキリンの仲間に、ほかより少し首の長いものが生まれました。より高い枝の葉を食べられて有利なので、首の長いキリンがより多く生き残ってきました。長い年月をかけてキリンの集団は首の長いものの割合が増えてゆきました。だから今私たちの見ているキリンはどれも首が長いのです。このように、ダーウィンが唱えた「自然選択説」の例として挙げられるキリンの首は有名ですね。ただ、自然選択説では、生物ごとの「進化しやすさ」の違いを説明することはできません。
今までの進化学では、突然変異は必ず起こる、生存や生殖に有利な突然変異は必ず残る、という仮定がありました。生物によって持つ遺伝情報は違いますが、その点はブラックボックスに入れたまま無視していたのです。しかし実は、有利な突然変異が起こるかどうかも、生まれた時に変異が集団中に広がっていくスピードも、生物によって大きく異なります。
田中先生の研究の狙いは、今まさに野外で起きている進化を理解すること。ダーウィンが唱えた自然選択説に、生物ごとに異なるDNA情報を結び付けて明らかにしようとしています。先生、進化を理解するってどういうことですか?
「系統樹を描くことで(この時こんな進化が起きただろう、と)推測するのではなくて、タイムマシンに乗っていくんだよ」と田中先生。タイムマシンとは、どういうことでしょう?例えば、爬虫類に羽ができて鳥類が生まれた、という進化の大イベント。地上に大型の捕食者がいたため、枝から飛び降りるときに、遠くに逃げられる羽を持つ方が有利だったのだと考えられています。もし、その瞬間のその生態系にタイムマシンで行けたら、そこでどの程度の自然選択が働いて、一世代あたりどの位の頻度で羽をつくる突然変異が集団に増えていった、という進化のプロセスをとらえることができます。
そんなふうに、あたかもタイムマシンに乗っているかのように、進化の現場をつぶさに実況するーそんなことができるのでしょうか。
ミヤマハタザオから、今起こっている進化をとらえる
6年前、先生は、現在進行形の進化を見る研究に適した植物を探し始めました。様々な大学や博物館・植物園の標本庫へ行き調べた結果、アブラナの一種であるミヤマハタザオが幅広い標高の場所で採られている、ということが分かりました。一種で多様な環境に適応していることに着目したのです。そこから過去に採集されたものが今もそこにいるか確かめたり、新しい生息場所を探すといった野外調査を始めました。

ミヤマハタザオは山のがけ崩れの後などに生える、アブラナ科の多年草。

ナズナの仲間であり可憐な白い花を咲かせる。
![]()
野外調査は苦労の連続。まず標本の採集地と同じ場所へ行くことさえもが一苦労でした。新しい標本にはGPSの緯度経度データがありますが、古い標本にはありません。標本の採集地が「仙丈ケ岳3000m」であれば、山頂付近を1時間くらい探したら見つかります。ところが「天竜村200m」のように標高の低い場所では、範囲が広くてどこをさすのか分かりにくく、一帯を探してもなかなか見つかりません。やっと採集地を見つけても、ミヤマハタザオがなくなっていることもあり得ます。通常、ミヤマハタザオはがけ崩れなどの荒れ地に生えるのですが、他の草に覆われて、姿を消してしまうこともあるのです。
やっと採集した種を栽培するのもまた一苦労です。田中先生は、これまでに知られていた方法で栽培をはじめましたが、1年目で実をつけたのはたったの3分の1。まず発芽しない、発芽しても開花をしない、開花しても種が出来ない、という課題を一つ一つ丁寧に克服して、独自の栽培方法を先生は築かれました。
ところが、そうして試行錯誤をしてきても、まだどうやっても発芽できない種が4集団あるそうです。「どうやっても発芽しないやつが、どうやったら発芽するのか非常に興味深いよね。種はぷりぷりなんだよ。すごい良い種なんだよ!なんでこの種で発芽しないのって感じなんだよね」。未だ眠り続けている種は、いったいどんな鍵を待っているのでしょうか。先生は今も様々な栽培方法、特に水攻め作戦を試しています。種をまく前に水に漬ける時間を、通常の倍以上にも長くしています。4つのうちいくつかはこれで発芽するかもしれない、と期待を寄せています。
ゆとりを活かして、進化中!
 このように苦労して得たミヤマハタザオからDNAを取り出し、生息地の異なるものの遺伝情報を比較します。その結果、先生は標高が違うと働きが異なる、光受容に関わる遺伝子をつきとめました。
このように苦労して得たミヤマハタザオからDNAを取り出し、生息地の異なるものの遺伝情報を比較します。その結果、先生は標高が違うと働きが異なる、光受容に関わる遺伝子をつきとめました。
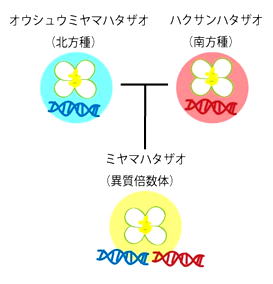
もともとミヤマハタザオは2つの親種に由来する雑種です(左図)。このような同じ遺伝情報を2セットと余分に持つ雑種は、身近なものではパンコムギなどが知られています。
ミヤマハタザオは北方種であるオウシュウミヤマハタザオと、南方種であるハクサンハタザオの雑種。両親由来のゲノムを重複してもち、同じ遺伝子が2つずつあるといえる。これを異質倍数体という。
生息地の標高が異なる集団の遺伝子をそれぞれ比べてみると、北方種由来の遺伝子は、いずれも親種とさしたる変化はありません。南方種由来の遺伝子だけ、標高間で大きく違っているし、祖先とも違っています(下図)。![]()
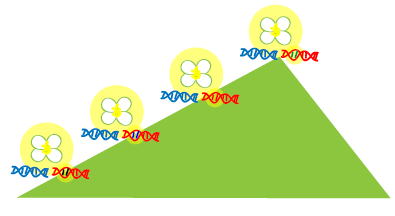
異質倍数化したミヤマハタザオには、1個の遺伝子は変化せずに本来の働きを保存しておいて、もう1個の遺伝子を標高に合わせて変化させる、「進化を可能にするゆとり」があることがわかりました。今までは「遺伝子重複説」という仮説に過ぎませんでしたが、これを今生きている生き物で見ることができたのです。
ミヤマハタザオの光受容に関わる遺伝子は、南方系由来の遺伝子のみが、生息地の標高に応じて変化していることが明らかになった。一方、北方系由来の遺伝子はほとんど変化していなかった。
![]() 田中先生の研究で明らかになったのは、片方の遺伝子をそのまま保存しておいて、もう一方を変化させるという進化の戦略です。異質倍数体が進化を加速する仕組みは、他にも考えられます。標高が高いところでは北方系由来の遺伝子が働き、低いところでは南方系由来の遺伝子が働くという、2つの遺伝子の使い分けもできそうです。「2セットの遺伝子の使い分け」という進化のプロセスもとらえたいと、先生は今研究に奮闘中です。
田中先生の研究で明らかになったのは、片方の遺伝子をそのまま保存しておいて、もう一方を変化させるという進化の戦略です。異質倍数体が進化を加速する仕組みは、他にも考えられます。標高が高いところでは北方系由来の遺伝子が働き、低いところでは南方系由来の遺伝子が働くという、2つの遺伝子の使い分けもできそうです。「2セットの遺伝子の使い分け」という進化のプロセスもとらえたいと、先生は今研究に奮闘中です。
【取材・構成・文 生物学類4年 遠藤愛】